
【论著】| 乳腺癌中线粒体功能障碍与CPT1A/ERK信号转导通路共同调节乳腺癌恶性行为的机制研究
2024-08-18 中国癌症杂志 中国癌症杂志
本研究旨在评估CPT1A过表达与线粒体动力学驱动的乳腺癌细胞转移之间的关系。
[摘要] 背景与目的:肉碱棕榈酰转移酶1A(carnitine palmitoyl transferase 1A,CPT1A)的高表达与乳腺癌患者预后较差相关,且能促进线粒体对脂肪酸的利用并最大限度地提高三磷酸腺苷(triphosphate,ATP)产量。然而,CPT1A在乳腺癌转移中的作用仍不清楚。本研究旨在探讨乳腺癌中线粒体功能障碍与CPT1A/细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)信号转导通路共同调节乳腺癌恶性行为的机制。方法:分别使用慢病毒系统和shRNA工具在人乳腺癌细胞系MDA-MB-231和MCF7中过表达或敲低CPT1A,将细胞分为NC组、CPT1A组和shCPT1A组。采用transwell实验检测细胞侵袭能力。采用蛋白质印迹法(Western blot)分析细胞中过氧化物酶体增殖物激活受体γ辅激活因子-1α(peroxisome proliferator-activated receptor γ coactivator-1α,PGC-1α)、ERK1/2和CPT1A蛋白的表达。采用线粒体红染色分析MDA-MB-231和MCF7细胞系的线粒体形态,并通过耗氧率分析线粒体呼吸能力。结果:与表达对照载体的细胞相比,CPT1A过表达导致MCF7细胞和MDA-MB-231细胞侵袭能力增强(P<0.05),shCPT1A敲低导致细胞侵袭能力降低(P<0.05)。与NC组相比,CPT1A组MDA-MB-231和MCF7细胞中线粒体分支长度显著变短(P<0.05),ERK1/2、PGC-1α表达显著增加(P<0.05),shCPT1A组MDA-MB-231和MCF7细胞中线粒体分支长度显著变长(P<0.05),ERK1/2、PGC-1α表达显著降低(P<0.05)。此外,与NC组相比,CPT1A组MDA-MB-231细胞基础和最大呼吸能力以及ATP产量显著增加 (P<0.05),而shCPT1A组MDA-MB-231细胞基础和最大呼吸能力以及ATP产量显著降低(P<0.05)。结论:CPT1A激活的ERK1/2-PGC-1α信号转导通路在线粒体分裂介导的乳腺癌细胞转移中发挥重要作用。
[关键词] 乳腺癌;线粒体;肉碱棕榈酰转移酶1A;细胞外信号调节激酶;恶性行为
[Abstract] Background and purpose: The overexpression of carnitine palmitoyl transferase 1A (CPT1A) is related to the poor prognosis of breast cancer, and it can promote the utilization of fatty acids by mitochondria and maximize triphosphate (ATP) production. However, the role of CPT1A in breast cancer metastasis is still unclear. This study aimed to explore the mechanism that mitochondrial dysfunction and CPT1A/extracellular signal-regulated kinase (ERK) signaling pathway in breast cancer jointly regulate the malignant behavior of breast cancer. Methods: The lentivirus system and shRNA tools were used to overexpress or knock down CPT1A in human breast cancer cell lines MDA-MB-231 and MCF7, and the cells were divided into NC group, CPT1A group and shCPT1A group. The invasion ability of cells was detected by transwell assay, and the protein expressions of peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α), ERK1/2 and CPT1A were analyzed by Western blot. The mitochondrial morphology of MDA-MB-231 and MCF7 cell lines was analyzed using mitochondrial red staining, and mitochondrial respiratory capacity was analyzed by oxygen consumption rate. Results: Compared with cells expressing control vector, overexpression of CPT1A resulted in enhanced invasion abilities of MCF7 cells and MDA-MB-231 cells (P<0.05), while knockdown of shCPT1A resulted in decreased invasion abilities of cells (P<0.05). Compared with NC group, the length of mitochondrial branches in MDA-MB-231 and MCF7 cells in CPT1A group was significantly shorter (P<0.05), and the expressions of ERK1/2 and PGC-1α increased significantly (P<0.05). In shCPT1A group, the length of mitochondrial branches in MDA-MB-231 and MCF7 cells increased significantly (P<0.05), and the expressions of ERK1/2 and PGC-1α decreased significantly (P<0.05). In addition, compared with NC group, the cellular basis, maximum respiratory capacity and ATP production of MDA-MB-231 in CPT1A group increased significantly (P<0.05), while the cellular basis, maximum respiratory capacity and ATP production of MDA-MB-231 in shCPT1A group decreased significantly (P<0.05). Conclusion: ERK1/2-PGC-1α activated by CPT1A Signaling pathway plays a key role in mitochondrial division mediated breast cancer cell metastasis.
[Key words] Breast cancer; Mitochondria; Carnitine palmitoyl transferase 1A; Extracellular signal-regulated kinase; Malignant behavior
乳腺癌是女性常见的恶性肿瘤之一,占新发癌症病例的30%,占所有癌症相关死亡的15%[1]。中国乳腺癌的发病率和死亡率持续上升[2]。尽管良性乳头状瘤的诊疗已取得重大进展,但对于晚期和转移性乳腺癌尚缺乏有效的治疗手段[3]。乳腺癌细胞能够迁移并侵入周围组织和淋巴管,随后到达靶器官[4]。线粒体构成了一个不断重组的动态网络,在几种基本的细胞功能中发挥关键作用,包括代谢调节、能量产生、钙稳态、活性氧产生和程序性细胞死亡。有研究[5]显示,线粒体动力学积极参与肿瘤发生和代谢可塑性,使癌细胞适应不利的环境条件,从而促进肿瘤进展。此外,肿瘤细胞转移对能量有很高的需求,肿瘤细胞转移方向的前沿是能量需求最强的区域[5]。碎片化有助于线粒体富集到肿瘤转移方向的前沿,从而更有效地为局部细胞骨架运动提供三磷酸腺苷(triphosphate,ATP)[6]。然而,线粒体动力学在转移中的作用尚未完全阐明。肉碱棕榈酰转移酶1A(carnitine palmitoyl transferase 1A,CPT1A)是一种调节脂肪酸转运和氧化的限速酶,位于线粒体外膜[7]。CPT1A功能障碍与线粒体功能受损相关,并会导致代谢疾病或紊乱,包括肝脂肪变性、阿尔茨海默病、肥胖和动脉粥样硬化[8]。CPT1A过表达会增加线粒体中的活性氧(reactive oxygen species,ROS),促进前列腺癌的进展[9]。近期研究[10]显示,CPT1A过表达与患者预后较差有关,并且随着乳腺癌的进展,CPT1A逐渐激活一系列下游途径促进酰基肉碱复合物的形成,然后该复合物通过穿梭系统穿过线粒体内膜促进线粒体对脂肪酸的利用并最大限度地提高ATP产量。然而,CPT1A在乳腺癌转移中的作用仍不清楚。本研究旨在评估CPT1A过表达与线粒体动力学驱动的乳腺癌细胞转移之间的关系。
1 材料和方法
1.1 细胞培养
人乳腺癌细胞系MDA-MB-231和MCF7购自美国典型培养物保藏中心。MDA-MB-231在 37 ℃、无CO2的条件下于含有10%胎牛血清(fetal bovine serum ,FBS,美国Life Technologies公司)的莱博维茨L-15培养基(美国Life Technologies公司)中培养。MCF7细胞在37 ℃、CO2体积分数为5%的条件下于含有10%FBS的DMEM培养基(美国Life Technologies公司)中培养。
1.2 细胞处理
为了考察CPT1A敲低对MDA-MB-231和MCF7细胞侵袭的影响,将细胞分为NC组和shCPT1A组。NC组和shCPT1A组细胞经NC和shCPT1A转染,测定MDA-MB-231和MCF7细胞侵袭。由上海吉玛制药技术有限公司使用pGPU6/GFP/Neo载体合成CPT1A-shRNAs (shCPT1A,正向序列为5’-CCGGACGGCGCGGGAGGAATAAATTCTCGAGAATTTATTCCTC CCGCGCCGTTTTTTG-3’,反向序列为5’-AATT CAAAAAACGGCGCGGGAGGAATAAATTCTC GAGAATTATTCCTCCCGCGCCGT-3’)和sh阴性对照RNA(NC,序列为5’-AACGCTTGGTAGTA CGGACTATGGACACGAATGGGCG-3’)。使用LipofectamineTM3000(美国Invitrogen公司)将质粒转染到MDA-MB-231和MCF7细胞中,采用蛋白质印迹法(Western blot)分析确认是否成功敲低了CPT1A基因。
为了考察CPT1A过表达对MDA-MB-231和MCF7细胞侵袭能力的影响,将细胞分为NC组和CPT1A组。由上海吉玛制药技术有限公司使用以下引物构建pIRES2-CPT1A质粒:正向引物序列为5’-CACCGTTCCTCAGAATTTATCCAGT-3’,反向引物序列为5’-AAACACTGGATAAATT CTGAGGAAC-3’。引物用于扩增全长人CPT1A编码序列,通过Nhe Ⅰ和Sac Ⅰ位点将其克隆到pIRES2-AcGFP载体(美国Clontech公司)中。然后,使用LipofectamineTM3000试剂将质粒瞬时转染到MDA-MB-231和MCF7细胞中,采用Western blot分析确认是否成功过表达了CPT1A基因。
为了进一步证实乳腺癌细胞迁移与线粒体分裂有关,将CPT1A过表达MDA-MB-231细胞分为二甲基亚砜(dimethyl sulfoxide,DMSO)组、线粒体分裂抑制剂Mdivi-1组和线粒体融合启动子M1组。将Mdivi-1(上海睿铂赛生物科技有限公司)以25 mmol/L的储备浓度溶解在DMSO中,并用培养基稀释以达到25 μmol/L的最终浓度。将M1(美国Sigma公司)以5 mmol/L的浓度溶解在DMSO中,并用培养液稀释以达到5 μmol/L的最终浓度。
为了研究CPT1A诱导的过氧化物酶体增殖物激活受体γ辅激活因子-1α(peroxisome proliferator-activated receptor γ coactivator-1α,PGC-1α)的激活是否依赖于细胞外信号调节激酶1/2(extra cellular regulated kinase1/2,ERK1/2)的磷酸化,用不同剂量的ERK1/2通路抑制剂PD0325901处理稳定过表达CPT1A的MDA-MB-231细胞8 h。将PD0325901(美国Selleckchem公司)以1 mmol/L的储备浓度重悬于DMSO中,并用培养基稀释到50、100和200 nmol/L。
1.3 蛋白质分析
使用RIPA裂解缓冲液(上海碧云天生物技术股份有限公司)制备全细胞裂解物或线粒体裂解物,并通过十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecylsulphate polyacrylamide gel electrophoresis,SDS-PAGE)解析等量的蛋白质(从10 μg到30 μg)。将蛋白质转移到PVDF膜(美国Millipore公司)上,并在4 ℃下与PGC-1α(1∶1 000,美国Cell Signaling Technology公司)、ERK1/2(1∶1 000,美国Cell Signaling Technology公司)、CPT1A(1∶1 000,美国Cell Signaling Technology公司)、阴离子通道蛋白(voltage dependent anion channel,VDAC, 1∶1 000,美国Cell Signaling Technology公司)、DNA甲基转移酶1(DNA methyltransferase 1,DNMT1,1∶1 000,美国Cell Signaling Technology公司)和GAPDH(1∶2 000,美国Cell Signaling Technology公司)一抗共同温育过夜。第2天,在室温下与辣根过氧化物酶标记的二抗(美国Cell Signaling Technology公司)共同温育1 h后,用增强化学发光试剂盒(南京诺唯赞生物科技股份有限公司)观察目标蛋白。所有实验至少进行3次。
1.4 采用transwell实验检测细胞侵袭能力
将处理后的细胞以2×104个/孔接种在预涂有100 μL 1 μg/μL matrigel基质(美国BD Biosciences公司)的transwell小室(美国Corning公司)中用于侵袭测定。分别将无血清培养液和完全培养液添加到上室和下室。温育24 h后,侵袭的细胞用4%多聚甲醛溶液固定,用1%结晶紫染色液染色,在显微镜下拍照。所有实验至少进行3次。
1.5 线粒体分支长度的量化
细胞用MitoTracker Red(美国Thermo Fisher公司)染色,并使用AX10显微镜(德国Carl Zeiss公司)成像。然后使用ImageJ和Mito形态学Marco Plug-in分析图像。所有实验至少进行3次。
1.6 线粒体的耗氧率分析
海马XFe96分析仪(美国Agilent Technologies公司)用于测定线粒体呼吸能力。将2×104个细胞接种于XF细胞微孔板中并在37 ℃下培养24 h。然后在测定前1 h将细胞与海马XF基础培养基(美国Agilent Technologies公司)温育。在测定过程中的指定时间点,将1 μmol/L寡霉素A、0.5 μmol/L FCCP和0.5 μmol/L鱼藤酮/抗霉素A(美国Agilent Technologies公司)按顺序注射到每个指定的孔中,以测量耗氧率。所有实验至少进行3次。
1.7 线粒体转录物定量
使用TRIzol(美国Invitrogen公司)提取全细胞RNA,并使用PrimeScript RT试剂盒(日本Takara Bio公司)反转录。然后用TaqMan基因表达测定线粒体DNA转录定量测定法测定线粒体转录物,按照制造商的说明使用18 S rRNA作为输入对照。所有实验至少进行3次。
1.8 统计学处理
使用Prism 9.0软件对数据进行统计分析。数据描述以x±s表示。两组或两组以上的比较用Student t检验或ANOVA检验,多组间两两比较采用LSD事后检验。采用双侧检验,P<0.05为差异有统计学意义。利用GEPIA数据库(http://gepia. cancer-pku.cn/)中乳腺癌病例资料分析正常乳腺组织及不同分期乳腺癌组织中CPT1A mRNA的表达,同时构建高CPT1A mRNA表达和低CPT1A mRNA表达乳腺癌患者的Kaplan-Meier图。
2 结 果
2.1 CPT1A过表达与乳腺癌转移相关
分析GEPIA数据库中乳腺癌病例资料的结果显示,在乳腺癌组织中观察到较高的CPT1A RNA表达水平(P<0.05),并且CPT1A表达随组织学分期的升高有增加趋势(图1A、1B)。CPT1A表达水平升高预示着乳腺癌患者预后更差(图1C)。上述发现提示在MCF7和MDA-MB-231乳腺癌细胞中稳定过表达CPT1A(图1D、1E)后,与表达对照载体的细胞相比,CPT1A的过表达导致MCF7细胞和MDA-MB-231细胞侵袭能力增强(P<0.05,图1F、1G)。用shRNA敲除MCF7和MDA-MB-231乳腺癌细胞中CPT1A的表达(图1H、1I),与表达对照载体的细胞相比,shCPT1A敲低导致MCF7细胞和MDA-MB-231细胞侵袭能力降低(P<0.05,图1J、1K)。
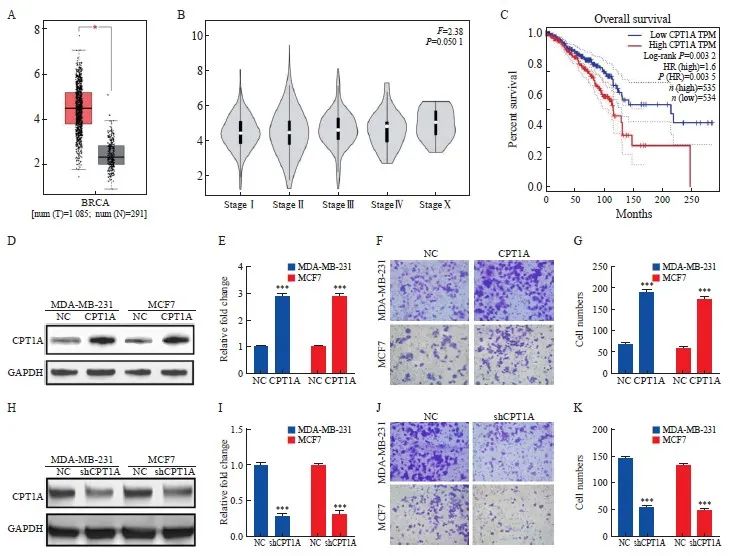
图1 乳腺癌组织中CPT1A的异常表达促进肿瘤转移
Fig. 1 Abnormal expression of CPT1A in breast cancer tissue promotes tumor metastasis
A, B: The expression of CPT1A mRNA in breast cancer tissues (n=1 085), normal breast tissues (n=291) and breast cancer tissues with different stages was analyzed by GEPIA. C: Using TCGA GEPIA to generate Kaplan-Meier diagram of breast cancer patients. The data of 1 069 breast cancer patients were divided into two groups according to the median percentile of CPT1A expression. D, E: Western blot representation and quantitative analysis of the expression level of CPT1A after overexpression. F, G: Transwell were used to determine the invasive ability of breast cancer cells expressing CPT1A or negative control vector. H, I: Western blot representation and quantitative analysis of the expression level of CPT1A after shRNA knockdown. J, K: Transwell were used to determine the invasive ability of breast cancer cells or negative control cells after CPT1A knock-down. *: P<0.05, compared with normal breast tissues; ***: P<0.001, compared with NC group.
2.2 CPT1A诱导的线粒体分裂是乳腺癌细胞侵袭所必需的
与NC组相比,CPT1A组MDA-MB-231和MCF7细胞中线粒体分支长度显著变短(P<0.05),shCPT1A组MDA-MB-231和MCF7细胞中线粒体分支长度显著变长(P<0.05,图2A~2D)。进一步分析线粒体网络动力学是否是乳腺癌细胞侵袭的原因。与用DMSO处理的细胞相比,在Mdivi-1和M-1处理的CPT1A过表达的MDA-MB-231细胞中均观察到线粒体分支长度显著变长(P<0.05),并且细胞侵袭能力显著降低(P<0.05,图2E~2H)。
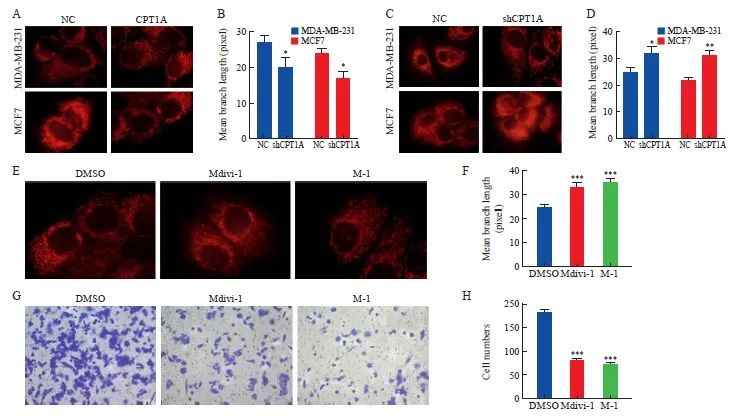
图2 CPT1A通过调节线粒体分裂促进乳腺癌细胞转移
Fig. 2 CPT1A promotes breast cancer cell metastasis by regulating mitochondrial division
A, B: Mitotracker living cell mitochondrial imaging and average mitochondrial branch length of cells overexpressing CPT1A or negative control. C, D: Mitotracker living cell mitochondrial imaging and average mitochondrial branch length of shRNA knock-down or negative control cells. E, F: Mitotracker mitochondrial imaging and average mitochondrial branching length of CPT1A overexpressing MDA-MB-231 cells treated with Mdivi-1 and M-1. G, H: Transwell was used to determine the invasive ability of CPT1A cells overexpressed MDA-MB-231 treated with Mdivi-1 and M-1. *: P<0.05, compared with NC group; **: P<0.01, compared with NC group; ***: P<0.001, compared with DMSO group.
2.3 CPT1A通过ERK1/2-PGC-1α途径促进线粒体分裂
为了进一步阐明CPT1A如何诱导线粒体分裂,进行了蛋白质抗体阵列分析,结果显示,ERK1/2表达明显增加(图3A)。Western blot分析显示,与NC组相比,CPT1A组MDA-MB-231细胞中ERK1/2表达和PGC-1α表达显著增加(P <0.05,图3B、3C)。为了研究CPT1A诱导的PGC-1α的激活是否依赖于ERK1/2,用不同剂量的ERK1/2通路抑制剂PD0325901处理稳定过表达CPT1A的MDA-MB-231细胞8 h。随着PD0325901浓度的增加,在MDA-MB-231细胞中观察到ERK1/2表达和PGC-1α表达均呈剂量依赖性抑制(P <0.05,图3D、3E)。
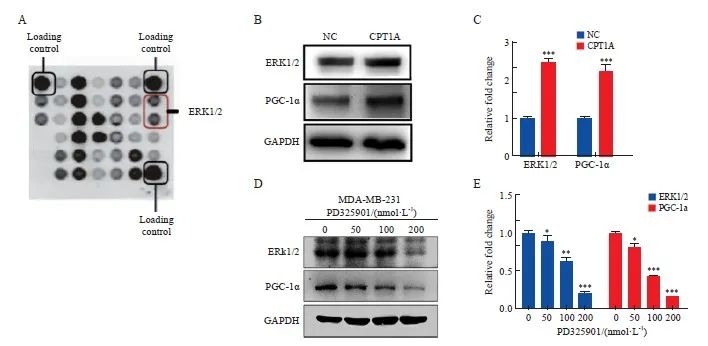
图3 CPT1A通过ERK1/2激活PGC-1α
Fig. 3 CPT1A activates PGC-1α through ERK1/2
A: Antibody array scanning of MDA-MB-231 cells overexpressing CPT1A. B, C: Western blot analysis and quantitative analysis of ERK1/2-PGC-1α expression in MDA-MB-231 cells over-expressing CPT1A. D, E: Western blot analysis and quantitative analysis of ERK1/2-PGC-1α expression in MDA-MB-231 cells treated with different doses of PD325901 for 8 h. *: P <0.05, compared with 0 nmol/L group; **: P <0.01, compared with NC group or 0 nmol/L group; ***: P <0.001, compared with NC group or 0 nmol/L group.
2.4 CPT1A促进线粒体氧化磷酸化
海马耗氧率(oxygen consumption rate,OCR)分析显示,与NC组相比,CPT1A组MDA-MB-231细胞基础和最大呼吸能力以及ATP产量显著增加(P<0.05),而shCPT1A组MDA-MB-231细胞基础和最大呼吸能力以及ATP产量显著降低(P<0.05,图4A~4D)。此外,Western blot分析显示,与NC组相比,CPT1A组MDA-MB-231细胞中PGC-1α蛋白表达和线粒体DNMT1表达均显著增加(P<0.05),而shCPT1A组MDA-MB-231细胞中PGC-1α和线粒体DNMT1表达均显著降低(P<0.05,图4E、4F)。复合体Ⅰ还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinamide adenine dinucleotide,NADH)对许多需氧生物的呼吸至关重要,其中7个疏水核心亚单位(命名为ND1~ND6和ND4L)由线粒体基因组编码。使用TaqMan基因表达分析来测量线粒体DNA转录,可量化ND1~ND6和ND4L的基因表达水平。与NC组相比,除ND6之外的ND1、ND2、ND3、ND4、ND4L和ND5亚单位在CPT1A组MDA-MB-231细胞中的表达水平均显著增加(P<0.05,图4G)。
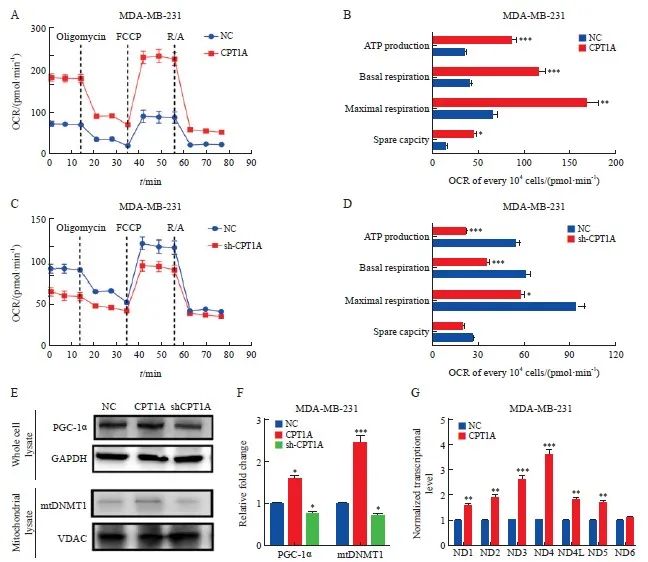
图4 CPT1A促进线粒体氧化磷酸化
Fig. 4 CPT1A promotes mitochondrial oxidative phosphorylation
A, B: Real-time OCR tracing of MDA-MB-231 cells overexpressing CPT1A and negative control and statistics of respiratory parameters. C, D: Real-time OCR tracing of MDA-MB-231 cells with CPT1A knockdown and negative control and statistics of respiratory parameters. E, F: Western blot analysis of the expression and quantitative analysis of PGC-1α and mitochondrial DNMT1 in MDA-MB-231 cells that overexpressed CPT1A and knocked down CPT1A. G: The expression level of mitochondrial DNA coding gene was measured by Taqman assay, and the data was standardized to 18s rRNA. *: P<0.05, compared with NC group; **: P<0.01, compared with NC group; ***: P<0.001, compared with NC group.
3 讨 论
本研究证明了CPT1A在线粒体分裂介导的乳腺癌细胞转移中的作用,并发现CPT1A在乳腺癌组织中呈高表达,CPT1A的高转录水平预示着患者预后不良。CPT1A通过激活ERK1/2-PGC-1α信号转导通路促进线粒体分裂,从而驱动乳腺癌细胞侵袭,可能是CPT1A诱导乳腺癌转移的新机制。
线粒体主要负责有氧能量的产生,线粒体的三羧酸循环和氧化磷酸化(oxidative phosphorylation,OXPHOS)是NADH和ATP的主要来源[11]。线粒体由单个连续的线粒体网状结构相互连接,其连接程度决定了线粒体融合水平,而线粒体分裂导致整体尺寸更小的片段化线粒体。有研究[12-13]显示,线粒体动力学可能参与介导心肌病、神经退行性疾病中的细胞活动。癌细胞往往呈现碎片化的线粒体[14]。然而,线粒体分裂如何以及在何种程度上导致肿瘤恶性方面仍有待确定。本研究发现,CPT1A参与介导MDA-MB-231和MCF7细胞系的线粒体形态变化,CPT1A过表达导致线粒体分支长度显著变短,CPT1A敲低导致线粒体分支长度显著变长。本研究利用线粒体分裂抑制剂Mdivi-1和线粒体融合启动子M1共同证明了线粒体分支长度变长导致MDA-MB-231和MCF7细胞侵袭能力显著降低,表明碎片化的线粒体有利于乳腺癌细胞转移。
有研究[15]表明,线粒体融合可能提高呼吸效率,mtOXPHOS产生ATP,而线粒体分裂在OXPHOS中发挥抑制作用。然而,这些结论是通过剥夺关键的线粒体融合介体MFN1/2和OPA1来实现的,这些介体不仅在线粒体形态调节中发挥作用,而且在维持泛醌水平和线粒体嵴结构方面发挥作用[16]。上述研究表明,除了调节线粒体融合,OPA1和MFN1/2在呼吸链超级复合物组装中还有许多其他重要功能。另外,线粒体裂变能够促进有氧呼吸。乳腺癌干细胞表现出较高的线粒体氧化磷酸化水平,这对于富集耐化疗的乳腺癌干细胞至关重要[17]。PGC-1α作为能量代谢的关键调节剂,其通过介导线粒体生物发生和OXPHOS促进癌细胞转移。研究显示,PGC-1α依赖性分裂在维持乳腺上皮干细胞样细胞的干细胞性、脑肿瘤起始细胞的致瘤潜力和乳腺癌干细胞信号转导中发挥作用[18]。用Mdivi-1抑制PGC-1α激活能够显著降低线粒体呼吸能力[19]。更重要的是,PGC-1α介导的线粒体分裂激活能够促进线粒体自噬,并改善老年苍蝇的线粒体呼吸[20]。本研究结果显示,CPT1A诱导的PGC-1α依赖性线粒体分裂可增强线粒体基础和最大呼吸能力以及ATP产量,与上述研究结果一致。上述研究表明,线粒体分裂是线粒体氧化磷酸化的关键。
为了进一步阐明CPT1A如何诱导线粒体分裂,本研究进行了蛋白质抗体阵列分析。与对照相比,过表达CPT1A的MCF7细胞中ERK1/2表达显著增加。ERK1/2是促分裂原活化的蛋白激酶家族的一员,参与细胞分裂和生长、信号转导、增殖以及其他生理功能[21]。有研究[22]表明,激活ERK1/2途径能够促进线粒体ATP合成,诱导肝癌细胞生长。值得注意的是,PGC-1α激活与ERK1/2途径密切相关[23-24]。本研究发现,CPT1A在MDA-MB-231细胞中的过表达会导致PGC-1α表达增加,推测CPT1A过表达通过诱导ERK1/2磷酸化以激活PGC-1α。为了研究CPT1A诱导的PGC-1α的激活是否依赖于ERK1/2磷酸化,用不同剂量的ERK1/2磷酸化抑制剂PD0325901处理稳定过表达CPT1A的MDA-MB-231细胞8 h。随着PD0325901浓度的增加,在MDA-MB-231细胞中观察到ERK1/2表达剂量依赖性抑制,并且PGC-1α逐渐降低。上述结果表明,CPT1A通过激活PGC-1α促进线粒体分裂。PGC-1α的上调可能驱动DNMT1分布到线粒体基因组,并导致线粒体DNA甲基化,从而导致线粒体转录物的表达上调[25]。本研究显示,乳腺癌细胞中PGC-1α上调导致了mtDNA甲基化增强和线粒体基因组编码的NADH亚单位转录增加,与上述研究结果一致。上述研究提示CPT1A通过调节线粒体降解和生物发生来促进线粒体质量控制。
本研究揭示了CPT1A介导的ERK1/2-PGC-1α信号转导通路在线粒体分裂中的关键作用,并且线粒体分裂又导致了乳腺癌细胞迁移。线粒体分裂的遗传和药理学抑制降低了乳腺癌细胞的侵袭性和细胞呼吸。这些结果表明阻断CPT1A介导的线粒体质量控制可以作为转移性乳腺癌的治疗方法。然而,由于线粒体动力学调控机制复杂,无法排除线粒体动力学是否在PGC-1α调节中发挥作用。我们假设在CPT1A过表达的情况下,PGC-1α表达可能是线粒体分裂的反馈效应:线粒体分裂和线粒体吞噬导致线粒体质量减少,为了维持线粒体体内平衡,PGC-1α表达被激活以触发线粒体生物发生,从而实现更快的线粒体更新和更好的质量控制。因此,CPT1A调控PGC-1α的机制仍需要在后续的研究中进一步阐明。
利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:
姜丹:文章初稿撰写,实验设计及验证,数据整理,实验数据分析;
宋国庆:实验验证,数据整理,实验数据分析;
王晓丹:文章审阅与修订。
[参考文献]
[1]GIAQUINTO A N, SUNG H, MILLER K D, et al. Breast cancer statistics, 2022[J]. CA Cancer J Clin, 2022, 72(6): 524-541.
[2]XIA C F, DONG X S, LI H, et al. Cancer statistics in China and United States, 2022: profiles, trends, and determinants[J]. Chin Med J, 2022, 135(5): 584-590.
[3]GALIANDRO F, AGNES S, MOSCHETTA G, et al. Prognostic factors in patients with breast cancer liver metastases undergoing liver rep: systematic review and meta-analysis[J]. Cancers, 2022, 14(7): 1691.
[4]KOTECHA R, TONSE R, RUBENS M, et al. Systematic review and meta-analysis of breast cancer brain metastasis and primary tumor receptor expression discordance[J]. Neurooncol Adv, 2021, 3(1): vdab010.
[5]VONA R, MILEO A M, MATARRESE P. Microtubule-based mitochondrial dynamics as a valuable therapeutic target in cancer[J]. Cancers, 2021, 13(22): 5812.
[6]KUMAR H, GUPTA N V, JAIN R, et al. A review of biological targets and therapeutic approaches in the management of triple-negative breast cancer[J]. J Adv Res, 2023, 54: 271-292.
[7]JIANG N, XING B Z, PENG R, et al. Inhibition of Cpt1a alleviates oxidative stress-induced chondrocyte senescence via regulating mitochondrial dysfunction and activating mitophagy[J]. Mech Ageing Dev, 2022, 205: 111688.
[8]SAVUKAITYTĖ A, BARTNYKAITĖ A, BEKAMPYTĖ J, et al. DDIT4 downregulation by siRNA approach increases the activity of proteins regulating fatty acid metabolism upon aspirin treatment in human breast cancer cells[J]. Curr Issues Mol Biol, 2023, 45(6): 4665-4674.
[9]JOSHI M, KIM J, D’ALESSANDRO A, et al. CPT1A over-expression increases reactive oxygen species in the mitochondria and promotes antioxidant defenses in prostate cancer[J]. Cancers, 2020, 12(11): 3431.
[10]DAS M, GIANNOUDIS A, SHARMA V. The role of CPT1A as a biomarker of breast cancer progression: a bioinformatic approach[J]. Sci Rep, 2022, 12(1): 16441.
[11]ZHANG W, DU X F, LIU B, et al. Engineering supramolecular nanomedicine for targeted near infrared-triggered mitochondrial dysfunction to potentiate cisplatin for efficient chemophototherapy[J]. ACS Nano, 2022, 16(1): 1421-1435.
[12]FORTE M, SCHIRONE L, AMERI P, et al. The role of mitochondrial dynamics in cardiovascular diseases[J]. Br J Pharmacol, 2021, 178(10): 2060-2076.
[13]YOU Y, MURAOKA S, JEDRYCHOWSKI M P, et al. Human neural cell type-specificextracellular vesicle proteome defines disease-related molecules associated with activated astrocytes in Alzheimer’s disease brain[J]. J Extracell Vesicles, 2022, 11(1): e12183.
[14]ANSARI M I, BANO N, KAINAT K M, et al. Bisphenol A exposure induces metastatic aggression in low metastatic MCF-7 cells via PGC-1α mediated mitochondrial biogenesis and epithelial-mesenchymal plasticity[J]. Life Sci, 2022, 302: 120649.
[15]MIDDLETON P, VERGIS N. Mitochondrial dysfunction and liver disease: role, relevance, and potential for therapeutic modulation[J]. Therap Adv Gastroenterol, 2021, 14: 17562848211031394.
[16]LIU Y E, SHI Y F. Mitochondria as a target in cancer treatment[J]. MedComm, 2020, 1(2): 129-139.
[17]LEE K M, GILTNANE J M, BALKO J M, et al. MYC and MCL1 cooperatively promote chemotherapy-resistant breast cancer stem cells via regulation of mitochondrial oxidative phosphorylation[J]. Cell Metab, 2017, 26(4): 633-647. e7.
[18]PRAHARAJ P P, PATRO B S, BHUTIA S K. Dysregulation of mitophagy and mitochondrial homeostasis in cancer stem cells: novel mechanism for anti-cancer stem cell-targeted cancer therapy[J]. Br J Pharmacol, 2022, 179(22): 5015-5035.
[19]ZHANG L, SUN L, WANG L R, et al. Mitochondrial division inhibitor (mdivi-1) inhibits proliferation and epithelial-mesenchymal transition via the NF-κB pathway in thyroid cancer cells[J]. Toxicol In Vitro, 2023, 88: 105552.
[20]RANA A, OLIVEIRA M P, KHAMOUI A V, et al. Promoting Drp1-mediated mitochondrial fission in midlife prolongs healthy lifespan of drosophila melanogaster[J]. Nat Commun, 2017, 8(1): 448.
[21]HU N, CHEN X M, CHEN C C, et al. Exploring the role of esketamine in alleviating depressive symptoms in mice via the PGC-1α/irisin/ERK1/2 signaling pathway[J]. Sci Rep, 2023, 13(1): 16611.
[22]HUANG K Y, LIU Z Y, XIE Z L, et al. HIGD2A silencing impairs hepatocellular carcinoma growth via inhibiting mitochondrial function and the MAPK/ERK pathway[J]. J Transl Med, 2023, 21(1): 253.
[23]LIU J F, NING L. Protective role of emodin in rats with post-myocardial infarction heart failure and influence on extracellular signal-regulated kinase pathway[J]. Bioengineered, 2021, 12(2): 10246-10253.
[24]LAURIN K M, COUTU-BEAUDRY K, SALAZAR A, et al. Low expression of PGC-1β and other mitochondrial biogenesis modulators in melanoma is associated with growth arrest and the induction of an immunosuppressive gene expression program dependent on MEK and IRF-1[J]. Cancer Lett, 2022, 541: 215738.
[25]ASL E R, AMINI M, NAJAFI S, et al. Interplay between MAPK/ERK signaling pathway and microRNAs: a crucial mechanism regulating cancer cell metabolism and tumor progression[J]. Life Sci, 2021, 278: 119499.
作者:中国癌症杂志
版权声明:
本网站所有注明“来源:梅斯医学”或“来源:MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明“来源:梅斯医学”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言








#乳腺癌# #线粒体# #肉碱棕榈酰转移酶1A# #细胞外信号调节激酶# #恶性行为#
44