JACS:中科大梁好均团队开发基于CRISPR-dCas9的可编程转录调节器
2022-07-07 “生物世界”公众号 “生物世界”公众号
为了有效控制基因表达,已经开发了三种主要策略来招募效应域:1)直接将效应域与 dCas9 融合;2)将 dCas9 与多聚肽阵列融合,以通过蛋白质招募相同效应域的多个拷贝-蛋白质相互作用。
CRISPR-dCas9 控制特定靶点基因表达的能力提供了一种通用的“基因开关”工具,可用于研究基因和调控元件的功能,产生不同的细胞类型或状态,并提供合成基因电路设计的强大策略。
为了有效控制基因表达,已经开发了三种主要策略来招募效应域:1)直接将效应域与 dCas9 融合;2)将 dCas9 与多聚肽阵列融合,以通过蛋白质招募相同效应域的多个拷贝-蛋白质相互作用;3)用 RNA 适体修饰 sgRNA 以通过 RNA-蛋白质相互作用募集效应结构域。
然而,这些策略旨在提高基于 dCas9 的转录调控的效率,同时缺乏一个单独的节点来精确控制基于 dCas9 的系统,这限制了 CRISPR-dCas9 系统实现的复杂合成电路的构建。为此,已经引入了化学或光诱导模块来对 CRISPR-dCas9 系统进行外部控制,但是可以很容易地编程以感知细胞信号的内部控制方法仍然很少。因此,引入基因模块来实现 CRISPR-dCas9 系统的内部控制,实现复杂的合成基因电路势在必行。
2022年7月6日,中国科学技术大学梁好均教授课题组在 JACS 期刊发表了题为:Programmable Transcriptional Modulation with a Structured RNA-Mediated CRISPR-dCas9 Complex 的研究论文。
该研究设计了一个基于 dCas9 的转录调控系统,该系统由一个单独的 RNA 模块介导,该模块可以通过RNA-蛋白质相互作用募集多个转录效应结构域。作为一个单独的组件,RNA 模块充当 dCas9 和效应域之间的桥梁,因此可以设计为生成更复杂的合成电路,用于基于dCas9 的基因操作的内部控制。此外,与直接将 RNA 适体引入 sgRNA 不同,单独的 RNA 模块可以在不干扰 sgRNA 结构的情况下进行任意修饰。
使用这一新系统,进一步提高了基因激活的效率,通过引入与 dCas9 融合竞争分离 RNA 模块的蛋白质,生成了响应内源性 microRNA 的电路,并实现了细胞混合物中的癌细胞识别。这些结果表明,单独的 RNA 介导的 dCas9 系统对于基因调控是有效的,并且对于实施合成电路是灵活的。

研究团队首先设计了一个 RNA 介导的基于 dCas9 的转录调控系统,其中 dCas9 和效应结构域分别融合到两个通过桥 RNA 连接的 RNA 结合蛋白(图1a)。桥 RNA 包含两个 RNA 模块,通过非键合双基础屈指连接,可以通过与相应结合蛋白的相互作用将 dCas9 与效应域连接,从而调节靶基因表达。首先,团队选择了三个特征明确的 RNA序列,MS2、PP7 和 COM,它们分别被 MCP、PCP 和 COM 蛋白识别用于桥 RNA 的设计。这里使用了一个突变的 MCP,即 MCP(V29I),其中第29位的缬氨酸被异亮氨酸取代,以增强与 MS2 的相互作用。为了简化表达,将其表示为MCP。
此外,为了改善桥 RNA 与 dCas9 融合的可能结合并将更多效应域募集到靶基因,使用两个重复的 MS2 或 PP7 发夹来构建桥 RNA。考虑到这些多发夹桥 RNA 的稳定性可能会受到限制,因此在每个模块的两个重复序列之间插入了一个双链接头。值得注意的是,在桥 RNA 中使用了单个 COM 发夹,因为 COM 蛋白与单个 COM 发夹底部的双链 RNA,和双链接头的引入可能会破坏 COM RNA 和 COM 蛋白的相互作用。使用 NUPACK 分析了所有桥 RNA 的二级结构。在设计中,在不同转录激活系统中表现良好的三个激活域,VP64、P65-HSF1 和 VPR 用于基因激活。
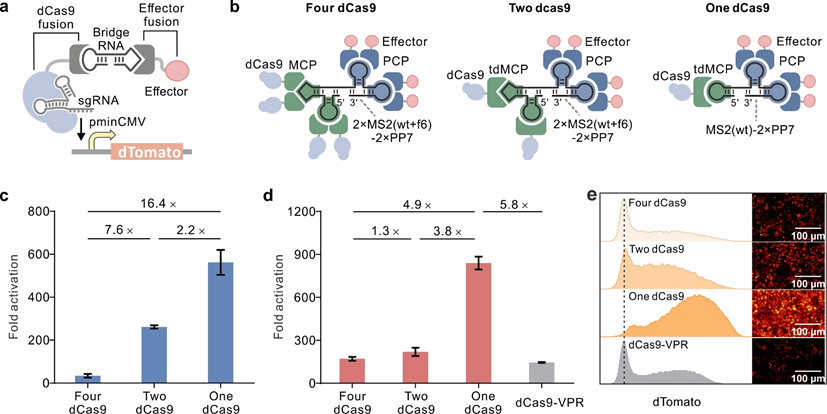
图1:桥 RNA 介导的基于 dCas9 的转录激活复合物的设计和优化
为了确定桥 RNA 介导的转录调节系统是否可以有效调节哺乳动物细胞中内源基因的表达,团队选择了先前研究中使用的几个基因(ASCL1、TNN、MIAT、NGN2、NeuroD1 和 ACTC1)(图2)。每个内源基因都被三个或四个 sgRNA 靶向。当 sgRNA 与桥 RNA 介导的复合物一起在 HEK293T 细胞中共表达时,观察到通过 qRT-PCR 测量的靶基因的 mRNA 水平显着增加(图2a)。此外,与传统使用的 dCas9-VPR 系统相比,桥 RNA 介导的转录激活系统使 NeuroD1 表达增加 6.1 倍,使 ACTC1 表达增加 2.9 倍(图2b)。此外,进一步的研究表明,具有高基础表达的基因在系统中更难激活,这与早期研究一致。

图2:用于 HEK293T 细胞内源基因激活的桥 RNA 介导的 dCas9 平台
为了生成仅在 microRNA 处于高水平时才允许基因激活的双反转系统,首先,引入了竞争对手来抑制基于 dCas9 的基因激活。在设计中,使用 tdMCP(V29I)蛋白与 dCas9-tdMCP 竞争结合桥 RNA MS2-2×PP7(图 3a)。竞争者破坏了桥 RNA 介导的转录激活复合物的形成,从而中断了报告基因的激活。为了促进竞争者与桥 RNA MS2-2×PP7 的结合,在 dCas9-tdMCP 的两个 MCP 中引入了一个单一突变,以削弱 dCas9 融合和桥 RNA MS2-2×PP7 之间的亲和力。dCas9 融合与 MCP 在不同位置发生突变,并产生了与野生型 MCP 的 dCas9 融合。然后,在桥 RNA 介导的系统中,在有或没有竞争者 tdMCP(V29I)的情况下,测试了五种 dCas9 融合的报告基因激活。结果表明,当单独使用五个 dCas9 融合时,报告基因的表达在没有竞争者的情况下增加了 65 到 646 倍。然而,在存在竞争者的情况下,荧光显着降低,证明了竞争者在与桥 RNA 结合方面的优势。为了获得更高的输出和更低的泄漏,dCas9-tdMCP(V29I)用于以下研究。接下来,为了抑制竞争对手的表达,将四个重复的 microRNA 完全互补的靶位点融合到竞争对手的 3'-非翻译区 (3'-UTR) 中,以构建 microRNA 传感器 (tdMCP(V29I)-TmiR- X)。一旦竞争对手被高水平的 microRNA 沉默,预计报告基因的高输出水平(图3a)。为了评估设计的可靠性,使用在 HEK293T 细胞中高度表达的 miR-17-5p、miR-20a 和 miR-106a来构建三个 microRNA 传感器。继续优化 dCas9 融合量和 dCas9 融合与竞争对手的比率,并观察到相对于缺乏 microRNA 靶位点的电路的输出高 52 到 118 倍,用于模拟低 microRNA 水平(图3b)。
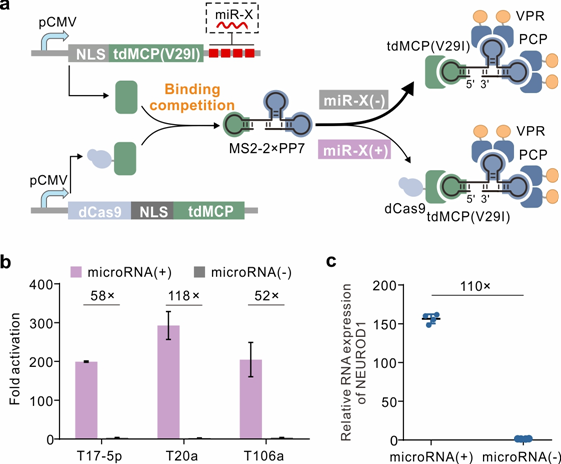
图3:响应 microRNA 操纵基因表达
癌细胞的识别和分离对于精确治疗具有重要意义。为了扩展用于细胞识别和分离的 microRNA 响应电路,通过将四个重复的 miR-21 靶标融合到竞争对手的 3 '-UTR (tdMCP(V29I)-T21)。MiR-21 在 HCT116 细胞中高表达,但在 HEK293T 细胞中很少表达,是分类这两种细胞类型的理想生物标志物。通过优化 dCas9 融合量和 dCas9 融合与竞争者的比例,实现了 HCT116 和 HEK293T 细胞之间 22 倍的荧光差异。电路性能的一个关键衡量标准是异质细胞群的特异性和选择性。
将电路输送到 HEK293T-EGFP 和 HCT116 细胞的细胞混合物中(图4a)。此外,在没有竞争者的情况下,由桥 RNA 介导的系统诱导的本体输出用作对照。在用完整电路转染的细胞混合物中,dTomato 高群体相对于对照强烈偏向 HCT116 细胞(图4b)。dTomato-high 细胞中 HCT116 的细胞丰度从 11.1% 增加到 92.2%(图4c),表明响应 miR-21 的桥 RNA 介导的系统是 HCT116 细胞识别和分离的有效平台。然而,由于 dCas9 融合和桥 RNA 之间的结合很少,HEK293T-EGFP 细胞中的电路显示出渗漏的背景。为了减少泄漏,可以通过将 microRNA 靶位点融合到 dCas9 融合或效应融合的 3'-UTR 中来引入由 HEK293T 高标记重新压制的传感器,以进行进一步研究。
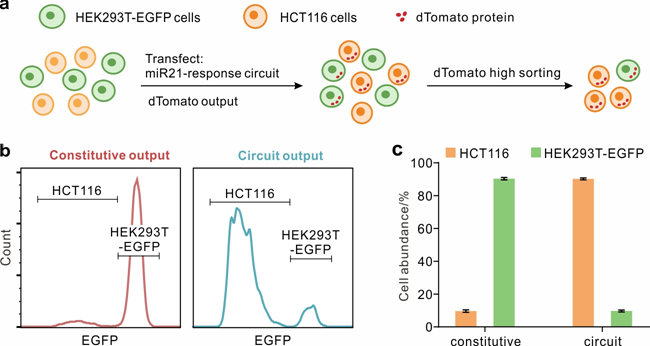
图4:细胞混合物中的 HCT116 细胞鉴定
中国科学技术大学高分子科学与工程系何苗博士和周翔博士后为论文共同第一作者,梁好均教授、姚东宝特任副研究员、周翔博士后为共同通讯作者。

梁好均,合肥微尺度物质科学国家研究中心研究员,中国科学技术大学高分子科学与工程系教授、博士生导师,2005年国家杰出青年基金获得者。梁好均教授已在 Nat. Rev. Methods Primers、PNAS、JACS、Angew. Chem. Int. Edit.、Adv. Mater.、ACS Nano、Nucleic. Acids. Res. 等高影响力学术期刊上发表论文两百余篇。梁好均教授于2022年2月11日13时10分在安徽合肥不幸离世,享年59岁。深切缅怀梁好均教授,感谢他在高分子模拟、DNA纳米材料以及合成生物学方面的突出贡献。
原始出处:
Miao He, Xiang Zhou, et al. Programmable Transcriptional Modulation with a Structured RNA-Mediated CRISPR-dCas9 Complex. . Am. Chem. Soc. 2022.
版权声明:
本网站所有注明“来源:梅斯医学”或“来源:MedSci原创”的文字、图片和音视频资料,版权均属于梅斯医学所有。非经授权,任何媒体、网站或个人不得转载,授权转载时须注明“来源:梅斯医学”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
在此留言







#中科大#
115
#转录#
147
#CRISPR#
116
#ACS#
104
#Cas9#
114